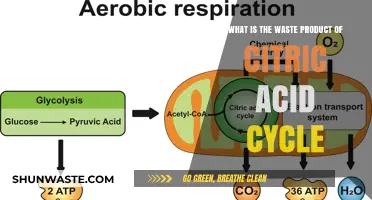
The electron transport chain (ETC) is a crucial process in cellular respiration, occurring in the inner mitochondrial membrane, where it generates ATP through oxidative phosphorylation. As electrons are passed along a series of protein complexes, energy is released, which is used to pump protons across the membrane, creating an electrochemical gradient. This gradient drives ATP synthesis via ATP synthase. However, the ETC also produces waste products, primarily in the form of water (H₂O). This water is generated when molecular oxygen (O₂) accepts the final electrons from the chain, combining with protons (H⁺) to form water, a benign and essential byproduct of aerobic respiration. Understanding this waste product is key to grasping the efficiency and sustainability of energy production in living organisms.
| Characteristics | Values |
|---|---|
| Waste Product | Carbon Dioxide (CO₂) and Water (H₂O) |
| Primary Source | Oxidative phosphorylation during cellular respiration |
| Location | Mitochondrial inner membrane (in eukaryotes) |
| Mechanism | Produced as electrons are passed through the electron transport chain, ultimately reducing molecular oxygen (O₂) |
| Chemical Reactions | CO₂ is produced during the Krebs cycle (Citric Acid Cycle), while H₂O is formed at the end of the electron transport chain when protons and electrons combine with oxygen |
| Role in Energy Production | Facilitates ATP synthesis via chemiosmosis and the proton gradient |
| Environmental Impact | CO₂ is a greenhouse gas, contributing to climate change when released in large quantities |
| Biological Significance | Essential for aerobic respiration in most organisms, providing energy for cellular processes |
| Alternative Pathways | In anaerobic respiration, waste products differ (e.g., lactic acid or ethanol) due to the absence of oxygen |
| Regulation | Controlled by factors like oxygen availability, substrate concentration, and metabolic demand |
Explore related products





What You'll Learn
- Carbon Dioxide Production: CO2 is released during the electron transport chain via oxidative phosphorylation
- Water Formation: Oxygen reduction at Complex IV forms water as a byproduct
- Superoxide Radicals: Incomplete oxygen reduction can generate harmful superoxide free radicals
- Proton Gradient Role: Protons pumped across the membrane are not waste but drive ATP synthesis
- Heat Dissipation: Some energy from the electron transport chain is lost as heat
![]()
Carbon Dioxide Production: CO2 is released during the electron transport chain via oxidative phosphorylation
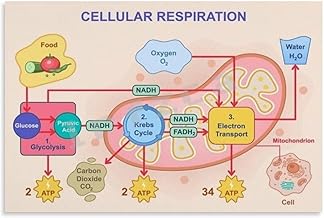
The electron transport chain (ETC) is a critical process in cellular respiration, where energy from nutrients is converted into adenosine triphosphate (ATP), the cell’s primary energy currency. During this process, electrons are passed through a series of protein complexes in the mitochondrial membrane, driving the production of ATP via oxidative phosphorylation. A key byproduct of this energy-generating pathway is carbon dioxide (CO₂), which is released as a waste product. This CO₂ originates from the oxidation of acetyl-CoA derived from glucose, fatty acids, or amino acids, highlighting the ETC’s role in breaking down organic molecules to release energy.
To understand CO₂ production in the ETC, consider the final steps of oxidative phosphorylation. Here, molecular oxygen (O₂) acts as the terminal electron acceptor, combining with electrons and hydrogen ions (H⁺) to form water (H₂O). However, the carbon atoms from the original fuel source (e.g., glucose) are not incorporated into water. Instead, they are released as CO₂ during the earlier stages of the citric acid cycle (Krebs cycle), which feeds into the ETC. For every molecule of glucose metabolized, six molecules of CO₂ are produced, demonstrating the direct link between substrate oxidation and CO₂ release.
From a practical standpoint, understanding CO₂ production in the ETC has implications for fields like exercise physiology and clinical diagnostics. During intense physical activity, muscle cells rely heavily on oxidative phosphorylation to meet energy demands, leading to increased CO₂ production and exhalation. This is why breath analysis, which measures CO₂ levels, is used to assess metabolic efficiency and aerobic fitness. For example, athletes may monitor their respiratory exchange ratio (RER), which compares CO₂ production to O₂ consumption, to optimize training regimens. An RER close to 1 indicates carbohydrate metabolism, while lower values suggest fat oxidation.
Comparatively, in anaerobic conditions or when the ETC is impaired, cells shift to glycolysis for energy production, generating lactic acid instead of CO₂. This highlights the ETC’s unique role in complete substrate oxidation and CO₂ release. In medical contexts, elevated blood CO₂ levels (hypercapnia) can signal respiratory or metabolic disorders, emphasizing the importance of the ETC in maintaining CO₂ homeostasis. Conversely, conditions like mitochondrial diseases, which disrupt ETC function, often result in reduced CO₂ production and energy deficits.
In conclusion, CO₂ production during the electron transport chain is a direct consequence of oxidative phosphorylation, reflecting the complete oxidation of carbon-based fuels. This process is not only central to energy metabolism but also serves as a diagnostic marker for metabolic health and efficiency. By examining CO₂ release, researchers and practitioners can gain insights into cellular function, energy utilization, and disease states, underscoring the ETC’s pivotal role in both physiology and pathology.
Unveiling the Truth: Evidence of Toxic Waste in Chemtrails
You may want to see also
Explore related products






![]()
Water Formation: Oxygen reduction at Complex IV forms water as a byproduct
The electron transport chain (ETC) is a critical process in cellular respiration, where electrons are passed through a series of protein complexes to generate ATP. At the final stage of this process, Complex IV, also known as cytochrome c oxidase, plays a pivotal role in reducing oxygen to form water. This reaction is not only essential for energy production but also highlights the elegance of biological systems in managing waste. The byproduct of this reduction reaction is water (H₂O), a molecule that is vital for life and easily recyclable within the cell and the environment.
Analyzing the mechanism, Complex IV catalyzes the transfer of four electrons to a single oxygen molecule, which simultaneously combines with four protons (H⁺) to produce two molecules of water. This process is highly efficient and ensures that oxygen, a potentially reactive molecule, is safely converted into a harmless and useful substance. The reaction can be summarized as: O₂ + 4e⁻ + 4H⁺ → 2H₂O. This equation underscores the importance of Complex IV in maintaining the balance of cellular redox reactions while minimizing toxic byproducts.
From a practical standpoint, understanding water formation at Complex IV has implications for medical research, particularly in studying mitochondrial disorders. Defects in Complex IV can lead to impaired oxygen reduction, resulting in reduced ATP production and the accumulation of reactive oxygen species (ROS). For instance, conditions like Leigh syndrome and Parkinson’s disease have been linked to Complex IV dysfunction. Researchers often focus on enhancing Complex IV activity or mitigating ROS damage through antioxidants like coenzyme Q10 or vitamin C, which can be supplemented at dosages of 100–200 mg/day for adults, depending on the severity of the condition.
Comparatively, water formation at Complex IV contrasts with other biological processes that produce waste, such as lactic acid fermentation or urea production in the liver. Unlike these byproducts, which require specific pathways for elimination, water is immediately reusable in cellular processes, such as hydration of macromolecules or as a medium for biochemical reactions. This efficiency makes the ETC’s waste product uniquely benign and highlights the evolutionary optimization of energy metabolism.
In conclusion, the formation of water at Complex IV is a testament to the precision of the electron transport chain. It not only completes the cycle of electron transfer but also ensures that the byproduct is beneficial rather than harmful. This process exemplifies how biological systems have evolved to maximize efficiency while minimizing waste, offering valuable insights for both scientific research and practical applications in health and medicine.
Waste Not, Gain Much: How Byproducts Boost Saccharomyces Survival
You may want to see also
Explore related products





![]()
Superoxide Radicals: Incomplete oxygen reduction can generate harmful superoxide free radicals
The electron transport chain (ETC), a critical process in cellular respiration, typically culminates in the reduction of oxygen to water. However, this process is not always flawless. Incomplete reduction of oxygen can lead to the formation of superoxide radicals (O₂⁻), highly reactive molecules that pose significant risks to cellular health. These radicals are a byproduct of electron "leakage" from the ETC, particularly at Complexes I and III, where electrons prematurely interact with molecular oxygen.
Superoxide radicals are inherently unstable due to their unpaired electron, making them highly reactive. This reactivity drives their involvement in oxidative stress, a condition where the balance between free radicals and antioxidants is disrupted. Oxidative stress can damage cellular components such as lipids, proteins, and DNA, contributing to aging, neurodegenerative diseases, and cancer. For instance, in mitochondria, superoxide radicals can initiate lipid peroxidation, a chain reaction that degrades cell membranes, impairing their function and integrity.
To mitigate the harmful effects of superoxide radicals, cells employ antioxidant defense mechanisms. Superoxide dismutase (SOD) is a key enzyme that catalyzes the conversion of superoxide radicals into hydrogen peroxide (H₂O₂), a less reactive species. However, if H₂O₂ is not further reduced to water by enzymes like catalase or glutathione peroxidase, it can still cause damage. This highlights the importance of a robust antioxidant network in maintaining cellular health.
Practical strategies to minimize superoxide radical production include lifestyle modifications that support mitochondrial function. Regular physical activity, for example, enhances mitochondrial efficiency, reducing electron leakage. Dietary interventions, such as consuming foods rich in antioxidants (e.g., berries, nuts, and leafy greens), can also bolster the body’s defense against oxidative stress. Additionally, avoiding environmental toxins like cigarette smoke and excessive alcohol, which increase free radical production, is crucial.
In clinical settings, supplementation with antioxidants like vitamin C, vitamin E, and coenzyme Q10 has been explored to counteract superoxide-induced damage. However, dosage and timing are critical; excessive antioxidant intake can disrupt cellular signaling pathways. For adults, a daily intake of 15–90 mg of vitamin C and 15–100 IU of vitamin E is generally recommended, though individual needs may vary based on age, health status, and environmental factors. Always consult a healthcare professional before starting any supplementation regimen.
In summary, superoxide radicals are a harmful byproduct of incomplete oxygen reduction in the electron transport chain, contributing to oxidative stress and cellular damage. Understanding their formation and impact underscores the importance of maintaining a balanced antioxidant system and adopting lifestyle habits that support mitochondrial health. By integrating dietary, behavioral, and, when necessary, clinical interventions, individuals can effectively minimize the risks associated with these reactive species.
How to Cancel Waste Connections Trash Service: A Step-by-Step Guide
You may want to see also
Explore related products






![]()
Proton Gradient Role: Protons pumped across the membrane are not waste but drive ATP synthesis
The electron transport chain (ETC) is often misunderstood as a process that generates waste products, but this perspective overlooks a critical aspect of cellular energy production. While it’s true that the ETC produces water as a byproduct of oxygen reduction, the protons (H⁺ ions) pumped across the mitochondrial or bacterial membrane are not waste. Instead, they are central to the cell’s energy currency: ATP synthesis. This proton gradient, established by the ETC, is a reservoir of potential energy that drives the final step of oxidative phosphorylation, proving that what might appear as waste is, in fact, a vital resource.
Consider the mechanism: as electrons pass through the ETC complexes, protons are actively transported from the mitochondrial matrix to the intermembrane space in eukaryotes, or across the plasma membrane in prokaryotes. This creates an electrochemical gradient, often referred to as the proton motive force (PMF). The PMF consists of both a pH difference (proton gradient) and an electrical potential. When protons flow back into the matrix through ATP synthase, this energy is harnessed to phosphorylate ADP to ATP. Without this gradient, the cell would lack the driving force needed for efficient energy production. Thus, the proton gradient is not a byproduct but a strategic intermediate in the cell’s energy economy.
To illustrate, imagine a hydroelectric dam. Water accumulates behind the dam, creating potential energy. When released, the water flows through turbines, generating electricity. Similarly, the proton gradient acts as the "dam," storing energy that is released through ATP synthase to produce ATP. This analogy underscores the elegance of cellular design, where every step is optimized for efficiency. In fact, the proton gradient is so crucial that cells invest significant energy in maintaining it, ensuring that even under stress, ATP production can continue.
Practical implications of this process are seen in medical and biochemical research. For instance, uncouplers like 2,4-dinitrophenol (DNP) disrupt the proton gradient by allowing protons to leak back into the matrix without driving ATP synthesis. While historically used as a weight-loss drug, DNP is highly toxic due to its ability to collapse the PMF, leading to overheating and metabolic crisis. Conversely, understanding the proton gradient has led to advancements in bioenergetics, such as developing proton pump inhibitors for treating acid reflux or studying mitochondrial diseases linked to ATP synthase dysfunction.
In conclusion, the proton gradient is a testament to the cell’s ingenuity in repurposing what might seem like waste into a cornerstone of energy production. By recognizing its role, we gain deeper insight into cellular metabolism and its vulnerabilities. Whether in research, medicine, or education, appreciating the proton gradient’s function shifts our perspective from waste to resource, highlighting the interconnectedness of biological systems.
Turning Human Waste into Clean Energy: Sustainable Solutions for a Greener Future
You may want to see also
Explore related products






![]()
Heat Dissipation: Some energy from the electron transport chain is lost as heat
The electron transport chain (ETC) is a highly efficient process, but it’s not perfect. As electrons move through the chain, pumping protons across the mitochondrial membrane, a significant portion of the energy released is not captured as ATP. Instead, it dissipates as heat. This phenomenon is a natural consequence of the thermodynamics governing biological systems. Unlike machines designed to minimize energy loss, living organisms prioritize flexibility and survival, allowing some energy to escape as thermal waste. This heat production is particularly noticeable in endothermic animals, where it contributes to maintaining body temperature, but it occurs in all cells to varying degrees.
From a practical standpoint, understanding heat dissipation in the ETC can inform strategies for optimizing cellular energy use. For instance, in athletes or individuals under physical stress, increased metabolic activity leads to higher heat production, which can be beneficial for warming muscles but may also require better hydration and cooling mechanisms to prevent overheating. Conversely, in cold environments, this natural heat generation becomes a survival advantage. However, excessive heat dissipation can also be a concern in certain medical conditions, such as mitochondrial disorders, where inefficient energy transfer exacerbates metabolic stress. Monitoring heat output in such cases could provide insights into disease progression or treatment efficacy.
Comparatively, the ETC’s heat dissipation contrasts with other cellular processes that aim to maximize energy conservation. For example, substrate-level phosphorylation in glycolysis directly produces ATP without significant heat loss. Yet, the ETC’s inefficiency serves a purpose: it prevents the overproduction of ATP, which could lead to cellular damage or metabolic imbalance. This trade-off highlights the evolutionary optimization of energy systems, where waste is not always a flaw but a feature. In this context, heat is not merely a byproduct but a regulated output that supports cellular and organismal homeostasis.
To illustrate, consider the brown adipose tissue (BAT) in mammals, specialized for non-shivering thermogenesis. Here, the uncoupling protein 1 (UCP1) disrupts the proton gradient, causing energy to be released as heat instead of driving ATP synthesis. This mechanism is particularly active in newborns and hibernating animals, where maintaining body temperature is critical. While this process bypasses the ETC’s typical efficiency, it underscores the adaptability of heat dissipation as a physiological tool. For those studying metabolic disorders or developing thermogenic therapies, targeting such mechanisms could offer novel interventions.
In conclusion, heat dissipation in the electron transport chain is not a mere inefficiency but a fundamental aspect of cellular energetics. It balances energy production with thermal regulation, supporting survival across diverse conditions. By recognizing its role, researchers and practitioners can better address metabolic challenges, from optimizing athletic performance to treating mitochondrial diseases. This perspective shifts the narrative from viewing heat as waste to appreciating it as a vital component of life’s energy economy.
Mastering RV Waste Tank Hauling: Tips for Safe and Efficient Transport
You may want to see also
Frequently asked questions
The primary waste product of the electron transport chain is water (H₂O), formed when molecular oxygen (O₂) combines with hydrogen ions (H⁺) and electrons at the end of the chain.
Water is produced during the final step of the electron transport chain when oxygen acts as the terminal electron acceptor, reacting with hydrogen ions and electrons to form H₂O.
No, water is the only waste product of the electron transport chain under aerobic conditions. However, in anaerobic conditions, alternative electron acceptors like sulfate or nitrate may produce different waste products.
Water is considered a waste product because it is a byproduct of the redox reactions in the electron transport chain and is not directly involved in energy production (ATP synthesis).
No, carbon dioxide is not a waste product of the electron transport chain. CO₂ is produced during the citric acid cycle (Krebs cycle), which occurs prior to the electron transport chain.