
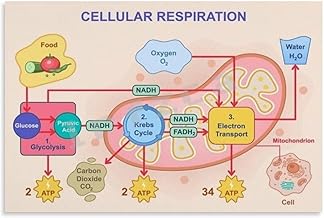
The electron transport chain (ETC) is a crucial process in cellular respiration, occurring in the mitochondria of eukaryotic cells, where it generates ATP, the cell's primary energy currency. As electrons are passed along a series of protein complexes in the ETC, energy is released and used to pump protons across the mitochondrial membrane, creating an electrochemical gradient. This gradient drives the synthesis of ATP through ATP synthase. However, not all energy derived from the electron transport chain is efficiently captured; a significant portion is lost as heat. Additionally, the final electron acceptor in the chain is molecular oxygen (O₂), which is reduced to form water (H₂O). While water is a benign and essential molecule for life, the heat generated during the process is considered a waste product of the electron transport chain, as it serves no direct biological function and is dissipated into the surrounding environment.
| Characteristics | Values |
|---|---|
| Name | Carbon Dioxide (CO₂) and Water (H₂O) |
| Primary Waste Product | CO₂ |
| Secondary Waste Product | H₂O |
| Source of CO₂ | Decarboxylation reactions in the Krebs cycle (Citric Acid Cycle) |
| Source of H₂O | Oxidation of NADH and FADH₂ during the electron transport chain |
| Role in Cellular Respiration | End products of oxidative phosphorylation |
| Transport Mechanism | CO₂ diffuses out of cells; H₂O remains in the cell or is excreted |
| Significance | Indicates complete oxidation of glucose and efficient ATP production |
| Environmental Impact | CO₂ is a greenhouse gas; H₂O is essential for life processes |
| Chemical Formula | CO₂: One carbon atom and two oxygen atoms; H₂O: Two hydrogen atoms and one oxygen atom |
| State at Room Temperature | CO₂: Gas; H₂O: Liquid |
Explore related products






What You'll Learn
- Carbon Dioxide Formation: CO2 is produced during decarboxylation steps in the citric acid cycle
- Water Production: Oxygen reduction at the end of the chain forms H2O
- Heat Generation: Energy not captured as ATP is released as thermal energy
- Superoxide Radicals: Incomplete oxygen reduction can produce reactive oxygen species
- Proton Gradient Dissipation: Leaky membranes may waste protons, reducing ATP synthesis efficiency
![]()
Carbon Dioxide Formation: CO2 is produced during decarboxylation steps in the citric acid cycle
The citric acid cycle, a central metabolic pathway, is not just a hub for energy production but also a significant source of carbon dioxide (CO2) in cellular respiration. During this cycle, CO2 is released in a process called decarboxylation, where a carboxyl group (COOH) is removed from intermediate molecules. This occurs twice in the cycle, specifically during the conversion of isocitrate to α-ketoglutarate and α-ketoglutarate to succinyl-CoA. Each decarboxylation step liberates one molecule of CO2, contributing to the overall production of waste products in cellular metabolism.
Analyzing the mechanism, decarboxylation is catalyzed by enzymes such as isocitrate dehydrogenase and α-ketoglutarate dehydrogenase. These enzymes facilitate the removal of CO2 by oxidizing the substrate, a process coupled with the reduction of NAD+ to NADH. This NADH is later utilized in the electron transport chain (ETC) to generate ATP, highlighting the interconnectedness of metabolic pathways. The CO2 produced here is a byproduct of breaking down carbon skeletons from glucose, fatty acids, or amino acids, emphasizing its role as a waste product rather than a functional molecule in this context.
From a practical standpoint, understanding CO2 formation in the citric acid cycle is crucial for fields like biochemistry and medicine. For instance, in diagnosing metabolic disorders, elevated CO2 levels in blood can indicate issues with cycle efficiency or substrate availability. Clinicians often measure serum CO2 as part of electrolyte panels, with normal ranges typically between 23–29 mEq/L. Deviations may prompt further investigation into mitochondrial function or dietary factors affecting substrate utilization. This underscores the importance of recognizing CO2 not just as waste but as a diagnostic marker.
Comparatively, while the electron transport chain primarily generates water and ATP, the citric acid cycle’s CO2 production serves a distinct purpose. Unlike water, which is a direct result of oxygen reduction, CO2 is a remnant of carbon metabolism. This distinction is vital in aerobic organisms, where CO2 must be efficiently expelled to prevent acidosis. For example, in humans, CO2 is transported via the bloodstream to the lungs and exhaled, a process regulated by respiratory mechanisms. This contrasts with anaerobic organisms, which lack such pathways and must manage CO2 accumulation differently.
In conclusion, CO2 formation during decarboxylation in the citric acid cycle is a critical yet often overlooked aspect of cellular respiration. It exemplifies how waste products are not merely discarded but are integral to metabolic regulation and diagnostic utility. By focusing on this specific process, we gain insights into the elegance of biochemical pathways and their broader implications in health and disease. Whether in a laboratory or clinical setting, appreciating this mechanism enhances our ability to interpret metabolic data and optimize physiological function.
Kurion's Innovative Plan to Tackle Fukushima's Tritium Nuclear Waste Crisis
You may want to see also
Explore related products






![]()
Water Production: Oxygen reduction at the end of the chain forms H2O
The electron transport chain (ETC) is a critical process in cellular respiration, where electrons are passed through a series of protein complexes to generate ATP. At the end of this chain, a fascinating and essential reaction occurs: the reduction of oxygen to form water (H₂O). This process is not merely a byproduct but a vital component of aerobic respiration, ensuring the sustainability of life in oxygen-dependent organisms.
The Chemistry Behind Water Formation
In the final step of the ETC, molecular oxygen (O₂) acts as the terminal electron acceptor. Electrons derived from NADH and FADH₂, along with protons (H⁺) from the mitochondrial matrix, combine with oxygen to produce water. The reaction can be simplified as follows: 4 electrons + 4 H⁺ + O₂ → 2 H₂O. This reduction process is catalyzed by cytochrome c oxidase, the last enzyme in the ETC. The efficiency of this reaction is remarkable; nearly 100% of the oxygen consumed is converted into water, minimizing the production of reactive oxygen species (ROS) that could damage cellular components.
Biological Significance of Water Production
Water formed during the ETC is more than just a waste product; it is a critical molecule for cellular function. In humans, this water contributes to the body’s overall hydration, though its direct impact is minimal compared to ingested water. However, in smaller organisms or in specific cellular contexts, this internally generated water can play a role in maintaining osmotic balance. For instance, in seeds during germination, water produced via the ETC aids in rehydration and metabolic activation. This highlights the dual role of the ETC: energy production and the synthesis of a life-sustaining molecule.
Practical Implications and Tips
Understanding water production in the ETC has practical applications, particularly in fields like bioenergetics and metabolic research. For example, measuring oxygen consumption and water production rates can assess mitochondrial efficiency, a key indicator of cellular health. In exercise physiology, optimizing ATP production via the ETC can enhance endurance, as efficient oxygen reduction ensures sustained energy output. To support this process, individuals can focus on consuming a diet rich in antioxidants (e.g., vitamins C and E) to protect the ETC from oxidative stress, ensuring optimal water formation and energy generation.
Comparative Perspective: Anaerobic vs. Aerobic Respiration
In contrast to aerobic respiration, anaerobic respiration does not produce water as a byproduct. Instead, it generates lactic acid or ethanol, depending on the organism. This comparison underscores the uniqueness of water formation in the ETC and its evolutionary advantage. Aerobic organisms, including humans, benefit from the clean, efficient energy production and the simultaneous synthesis of water, a molecule indispensable for life. This distinction also explains why aerobic respiration yields significantly more ATP (up to 36 molecules per glucose) compared to anaerobic pathways (only 2 ATP molecules).
By focusing on the specific process of oxygen reduction and water formation, we gain a deeper appreciation for the elegance and efficiency of the electron transport chain. This mechanism not only powers life but also ensures the production of a molecule as fundamental as water, reinforcing the interconnectedness of biological processes.
Efficient Residential Waste Pipe Installation Guide for California Homes
You may want to see also
Explore related products






![]()
Heat Generation: Energy not captured as ATP is released as thermal energy
The electron transport chain (ETC) is a highly efficient process, but it's not perfect. As electrons move through the chain, a significant portion of their energy is captured and used to generate ATP, the cell's primary energy currency. However, not all energy is harnessed in this way. A substantial amount of energy is lost as heat, a natural byproduct of the ETC's operation. This heat generation is a critical aspect of cellular metabolism, particularly in organisms that rely on non-shivering thermogenesis to maintain body temperature.
From an analytical perspective, the heat produced during the ETC can be understood through the lens of thermodynamics. The process involves a series of redox reactions where electrons are transferred from high-energy molecules to lower-energy ones. While some energy is conserved in the form of ATP through oxidative phosphorylation, the remainder is dissipated as thermal energy due to the inefficiency of energy transfer. This inefficiency is not a flaw but a feature, as it allows organisms to regulate temperature. For example, brown adipose tissue in mammals contains uncoupling proteins that deliberately increase heat production by decoupling the ETC from ATP synthesis, a mechanism essential for newborns and hibernating animals.
Instructively, understanding heat generation in the ETC has practical implications for health and medicine. For instance, therapies targeting brown adipose tissue activation are being explored to combat obesity and metabolic disorders. By increasing non-shivering thermogenesis, individuals can burn more calories without physical activity. A simple tip for boosting this process naturally includes exposure to mild cold, which stimulates brown fat activity. Additionally, certain dietary components like capsaicin (found in chili peppers) have been shown to enhance thermogenesis by increasing mitochondrial uncoupling.
Comparatively, heat generation in the ETC differs across species based on their metabolic needs. Ectotherms, such as reptiles, rely heavily on external heat sources and have less efficient ETC heat production. In contrast, endotherms, like mammals and birds, have evolved mechanisms to maximize heat generation, ensuring stable body temperatures in varying environments. This comparison highlights the adaptability of the ETC as both an energy-producing and heat-generating system, tailored to the survival needs of different organisms.
Finally, from a descriptive standpoint, the process of heat generation in the ETC is a vivid example of nature’s ingenuity. Imagine the mitochondria as tiny power plants within cells, where the flow of electrons not only fuels life but also warms it. This dual functionality is particularly evident in cold-adapted species, where the ETC’s role shifts from primarily energy production to heat generation. For instance, Arctic mammals like seals have a higher density of mitochondria in their muscles, enabling them to sustain both activity and warmth in frigid conditions. This interplay between energy capture and heat release underscores the ETC’s central role in balancing cellular and organismal needs.
Easy Steps to Install Waste Toner Bottle on Magicolor Printer
You may want to see also
Explore related products





![]()
Superoxide Radicals: Incomplete oxygen reduction can produce reactive oxygen species
The electron transport chain (ETC), a critical process in cellular respiration, typically culminates in the reduction of oxygen to water. However, under certain conditions, this process can be incomplete, leading to the formation of superoxide radicals (O₂⁻). These highly reactive molecules are a byproduct of electron leakage, primarily from Complexes I and III of the ETC. Unlike water, superoxide radicals are not inert; they are a subset of reactive oxygen species (ROS) that can initiate oxidative stress, damaging cellular components such as lipids, proteins, and DNA. This incomplete reduction is more likely to occur when the ETC is overburdened, such as during intense metabolic activity or in the presence of toxins that disrupt electron flow.
To understand the implications, consider the dosage of oxidative stress on cells. Even low levels of superoxide radicals can accumulate over time, particularly in post-mitotic cells like neurons and cardiomyocytes, which are less capable of regeneration. For instance, in aging individuals, the gradual buildup of ROS, including superoxide, is linked to degenerative diseases such as Alzheimer’s and cardiovascular disorders. Conversely, acute exposure to high levels of superoxide, as seen in conditions like ischemia-reperfusion injury, can cause immediate cellular damage. Practical tips to mitigate this include maintaining a balanced diet rich in antioxidants (e.g., vitamins C and E) and avoiding environmental stressors like cigarette smoke, which exacerbate ROS production.
From a comparative perspective, superoxide radicals differ from other ROS like hydrogen peroxide (H₂O₂) and hydroxyl radicals (·OH) in their reactivity and longevity. Superoxide is relatively stable and can diffuse across membranes, whereas hydroxyl radicals are short-lived but extremely reactive. The body employs enzymes like superoxide dismutase (SOD) to convert superoxide into less harmful H₂O₂, which is then neutralized by catalase or glutathione peroxidase. However, when SOD activity is overwhelmed, superoxide accumulates, tipping the balance toward oxidative damage. This highlights the importance of supporting endogenous antioxidant systems, especially in vulnerable populations such as the elderly or those with chronic illnesses.
Instructively, reducing superoxide production involves optimizing mitochondrial function. Regular physical activity, for example, enhances mitochondrial efficiency and increases antioxidant capacity, thereby lowering the risk of electron leakage. Conversely, excessive calorie intake or a sedentary lifestyle can impair mitochondrial health, promoting ROS generation. For individuals over 40, incorporating moderate aerobic exercise (e.g., 150 minutes per week) and strength training can be particularly beneficial. Additionally, supplements like coenzyme Q10 (CoQ10) or alpha-lipoic acid may support mitochondrial integrity, though their efficacy varies and should be discussed with a healthcare provider.
Finally, the analytical takeaway is that superoxide radicals are not merely waste products but markers of mitochondrial dysfunction. Their presence signals an imbalance between ROS production and antioxidant defenses, which, if unaddressed, can accelerate cellular aging and disease progression. By understanding the mechanisms of superoxide formation and implementing targeted interventions, individuals can proactively protect their cellular health. This underscores the need for a holistic approach to wellness, combining lifestyle modifications, dietary choices, and, when necessary, medical interventions to maintain the delicate equilibrium of cellular respiration.
Loose Bowels: Effective Waste Removal or Just a Quick Flush?
You may want to see also
Explore related products





![]()
Proton Gradient Dissipation: Leaky membranes may waste protons, reducing ATP synthesis efficiency
The electron transport chain (ETC) is a marvel of cellular efficiency, but even this finely tuned system can falter. One significant issue arises from leaky mitochondrial membranes, which allow protons to escape before they can be harnessed for ATP synthesis. This proton gradient dissipation is a silent saboteur, reducing the energy output of the cell. Imagine a hydroelectric dam with leaks—water (or in this case, protons) bypasses the turbines, generating less electricity. Similarly, when protons leak across the membrane, the electrochemical gradient collapses, and the ATP synthase enzyme cannot operate at full capacity.
To understand the impact, consider the numbers: under optimal conditions, the ETC can achieve a 40-50% efficiency in converting energy from electrons to ATP. However, proton leakage can slash this efficiency by up to 20%, depending on the severity of the leak. This is particularly problematic in cells with high energy demands, such as neurons and muscle cells. For instance, in aging cells, mitochondrial membranes often become more permeable, exacerbating this issue and contributing to age-related energy decline.
Addressing proton leakage requires a two-pronged approach. First, maintaining mitochondrial health is crucial. Antioxidants like Coenzyme Q10 (100-200 mg daily) and alpha-lipoic acid (300-600 mg daily) can help reduce oxidative damage to mitochondrial membranes. Second, compounds like SS-31, a mitochondrial-targeted peptide, show promise in stabilizing membranes and reducing proton leakage. While still in research stages, these interventions highlight the potential for targeted therapies to mitigate this inefficiency.
A comparative perspective reveals the elegance of nature’s solutions. Plants, for instance, have evolved mechanisms to minimize proton leakage in their chloroplasts, ensuring maximal energy capture during photosynthesis. By studying these adaptations, scientists can glean insights into improving mitochondrial efficiency in humans. For now, practical steps include adopting a diet rich in polyphenols (found in berries, nuts, and green tea) and engaging in regular exercise, both of which enhance mitochondrial function and reduce membrane permeability.
In conclusion, proton gradient dissipation due to leaky membranes is a critical yet underappreciated waste product of the electron transport chain. By understanding its mechanisms and implementing targeted strategies, we can potentially reclaim lost energy and improve cellular efficiency. Whether through dietary supplements, lifestyle changes, or future therapies, addressing this issue holds promise for combating age-related decline and enhancing overall vitality.
Did Republicans Vote to Allow Coal Waste Dumping in Rivers?
You may want to see also
Frequently asked questions
The primary waste product of the electron transport chain is water (H₂O), formed when molecular oxygen (O₂) combines with hydrogen ions (H⁺) and electrons at the end of the process.
Water is produced when oxygen acts as the final electron acceptor in the electron transport chain. Electrons and hydrogen ions combine with oxygen in a process catalyzed by the enzyme cytochrome oxidase, resulting in the formation of water.
No, water is the only waste product of the electron transport chain under aerobic conditions. However, in anaerobic respiration, alternative electron acceptors (e.g., sulfate or nitrate) are used, and different waste products (e.g., lactic acid or ethanol) are produced instead.































