
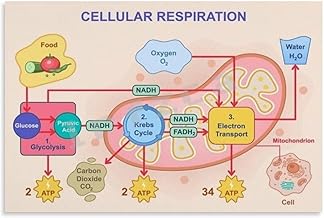
The electron transport chain (ETC) is a crucial process in cellular respiration, occurring in the inner mitochondrial membrane, where it generates ATP through oxidative phosphorylation. As electrons are passed along a series of protein complexes, energy is released, driving the production of ATP. However, this process also results in the formation of waste products, primarily in the form of water and carbon dioxide. Water is produced when molecular oxygen (O₂) accepts the final electrons from the ETC, combining with hydrogen ions (H⁺) to form H₂O. Additionally, carbon dioxide is generated earlier in the respiratory pathway, during the citric acid cycle, as a byproduct of the breakdown of acetyl-CoA derived from glucose. These waste products are essential indicators of the efficiency and completion of the electron transport chain and cellular respiration as a whole.
| Characteristics | Values |
|---|---|
| Primary Waste Products | Carbon Dioxide (CO₂), Water (H₂O) |
| Source of CO₂ | Decarboxylation reactions in the Krebs cycle (e.g., from pyruvate and α-ketoglutarate) |
| Source of H₂O | Oxygen (O₂) reduction at the end of the electron transport chain (ETC) |
| Role of O₂ | Final electron acceptor in the ETC, forming H₂O when combined with protons (H⁺) and electrons (e⁻) |
| Energy Efficiency | ~36-40 ATP molecules produced per glucose molecule, with CO₂ and H₂O as byproducts |
| Location in Cell | Mitochondrial matrix (Krebs cycle) and inner mitochondrial membrane (ETC) |
| Significance | CO₂ is excreted via respiration; H₂O is reused in cellular processes or excreted |
| Additional Byproducts | Heat (as a result of non-ATP-generating proton leaks across the membrane) |
| Environmental Impact | CO₂ is a greenhouse gas, contributing to climate change when released in large quantities |
| Biological Importance | Essential for energy production in aerobic organisms through oxidative phosphorylation |
Explore related products






What You'll Learn
- Carbon Dioxide Production: CO2 is released during decarboxylation steps in the citric acid cycle
- Water Formation: Oxygen is reduced to water at the end of the electron transport chain
- Heat Generation: Energy not captured in ATP is dissipated as thermal energy
- Superoxide Radicals: Incomplete oxygen reduction can produce reactive oxygen species (ROS)
- Proton Gradient Collapse: Protons leak back into the matrix, reducing ATP synthesis efficiency
![]()
Carbon Dioxide Production: CO2 is released during decarboxylation steps in the citric acid cycle
The citric acid cycle, a central metabolic pathway, is a powerhouse of energy production, but it also generates waste in the form of carbon dioxide (CO2). This process, known as decarboxylation, is a critical step in the cycle, where carbon atoms are removed from molecules, releasing CO2 as a byproduct. During the citric acid cycle, also referred to as the Krebs cycle or TCA cycle, two decarboxylation reactions occur, each resulting in the release of one molecule of CO2. These reactions are catalyzed by specific enzymes, ensuring the efficient removal of carbon dioxide from the intermediates.
The Decarboxylation Process:
In the first decarboxylation step, isocitrate dehydrogenase converts isocitrate to alpha-ketoglutarate, releasing CO2. This reaction is a crucial regulatory point in the cycle, as it is often the rate-limiting step. The second decarboxylation occurs when alpha-ketoglutarate dehydrogenase transforms alpha-ketoglutarate into succinyl-CoA, again producing CO2. These reactions are not only essential for the cycle's progression but also contribute to the overall carbon balance within the cell.
Implications and Significance:
The release of CO2 during these decarboxylation steps has several implications. Firstly, it highlights the citric acid cycle's role in carbon metabolism, where carbon atoms from nutrients are oxidized, ultimately forming CO2. This process is particularly important in aerobic organisms, as it links carbohydrate, fat, and protein metabolism to energy production. Secondly, the production of CO2 is a natural and necessary consequence of energy generation, emphasizing the delicate balance between energy yield and waste management within cellular processes.
From a practical perspective, understanding these decarboxylation reactions is vital in various fields. In biochemistry, it aids in comprehending metabolic disorders and designing targeted therapies. For instance, defects in the enzymes involved can lead to conditions like citrullinemia, causing ammonia to accumulate in the blood. In environmental science, the study of CO2 production in cellular respiration contributes to our understanding of carbon cycling and its impact on climate change.
In summary, the citric acid cycle's decarboxylation steps are not merely waste-producing reactions but integral components of energy metabolism. They showcase the intricate dance of molecules within cells, where energy extraction and waste generation are intricately linked. By examining these processes, scientists can unravel the complexities of metabolic pathways and their broader implications in health, disease, and environmental science. This knowledge is essential for developing strategies to optimize energy production and manage waste, both at the cellular and global levels.
Non-Biodegradable Waste: Environmental Impact and Sustainable Solutions
You may want to see also
Explore related products






![]()
Water Formation: Oxygen is reduced to water at the end of the electron transport chain
Oxygen's role in the electron transport chain culminates in a transformative process: its reduction to water. This final step, occurring at Complex IV (cytochrome c oxidase), is a cornerstone of aerobic respiration. Here, four electrons, funneled through the chain from NADH and FADH2, combine with four protons and molecular oxygen (O2) to produce two molecules of water (H2O). This reaction not only completes the electron transport chain but also highlights the elegance of biological systems in recycling and repurposing molecules.
From an analytical perspective, the reduction of oxygen to water is a redox reaction, where oxygen acts as the final electron acceptor. This process is highly efficient, ensuring that the energy released during electron transfer is captured in the form of ATP. The stoichiometry of the reaction is precise: for every molecule of oxygen consumed, two molecules of water are produced. This balance is critical for maintaining cellular homeostasis, as disruptions can lead to the formation of reactive oxygen species (ROS), which are harmful byproducts.
Instructively, understanding this process has practical implications in fields like medicine and biochemistry. For instance, drugs targeting Complex IV, such as cyanide, inhibit oxygen reduction, leading to cellular asphyxiation. Conversely, antioxidants that neutralize ROS are designed to mitigate damage caused by incomplete oxygen reduction. Researchers and clinicians can leverage this knowledge to develop therapies for conditions like mitochondrial diseases or ischemia-reperfusion injuries, where electron transport chain dysfunction plays a role.
Comparatively, the formation of water as a waste product contrasts with anaerobic respiration, where lactic acid or ethanol are produced instead. This distinction underscores the efficiency of aerobic respiration, which generates significantly more ATP per glucose molecule. Water, being a stable and inert molecule, is an ideal waste product, as it does not accumulate toxically within cells. This efficiency is why aerobic organisms dominate complex life forms, from insects to mammals.
Descriptively, the process is akin to a molecular assembly line, where oxygen is the final worker, accepting electrons and protons to create water. This metaphorical "assembly line" is powered by the proton gradient across the mitochondrial membrane, driving ATP synthesis. The end result—water—is not just a waste product but a testament to the precision and economy of cellular metabolism. By examining this step, we gain insight into the intricate dance of molecules that sustains life.
Cellular Waste Management: How Cells Remove and Recycle Waste Efficiently
You may want to see also
Explore related products






![]()
Heat Generation: Energy not captured in ATP is dissipated as thermal energy
The electron transport chain (ETC) is a highly efficient process, but it’s not perfect. As electrons move through the chain, their energy is gradually released and captured to form ATP, the cell's primary energy currency. However, not all energy is harnessed in this process. A significant portion—approximately 40-60%—escapes as thermal energy, contributing to the body’s heat production. This phenomenon is particularly vital in endothermic organisms, including humans, where heat generation helps maintain core body temperature. For instance, during cold exposure, the ETC ramps up activity in brown adipose tissue, intentionally producing heat to counteract the drop in external temperature.
From a practical standpoint, understanding this heat generation can inform strategies for managing metabolic health. For adults over 65, who often experience reduced thermoregulation, encouraging mild physical activity can stimulate the ETC and enhance heat production, aiding in warmth retention. Similarly, in infants, whose thermoregulatory systems are still developing, ensuring a warm environment is critical, as their ETC is less efficient at generating heat compared to adults. A simple tip: layering clothing rather than relying on a single thick garment allows for better heat retention by trapping air close to the skin.
Comparatively, this heat dissipation contrasts with the efficiency of ATP synthesis in processes like oxidative phosphorylation. While ATP production is a precise, stepwise mechanism, heat generation is more diffuse, occurring as a byproduct of proton leakage across the mitochondrial membrane. This inefficiency isn’t a flaw but a feature, especially in hibernating animals or during fever, where controlled heat production is essential for survival. For example, during hibernation, the ETC in bears shifts to prioritize heat over ATP, allowing them to maintain body temperature without expending excessive energy on movement.
To optimize energy use and minimize unnecessary heat generation, certain dietary and lifestyle adjustments can be made. Consuming a balanced diet rich in coenzyme Q10 (found in fatty fish and nuts) supports efficient ETC function, reducing excess heat production. Conversely, in athletes or those in cold climates, intentionally increasing metabolic rate through high-intensity interval training (HIIT) can enhance heat generation, improving cold tolerance. A cautionary note: excessive heat production without adequate hydration can lead to overheating, so maintaining fluid intake is crucial, especially during prolonged physical activity or in hot environments.
In conclusion, heat generation from the ETC is a natural, often beneficial byproduct of energy metabolism. By recognizing its role, individuals can tailor their behaviors—whether through diet, exercise, or environmental adjustments—to harness or mitigate this thermal energy as needed. For instance, a 30-minute session of HIIT three times a week can boost heat production in cold-sensitive individuals, while staying hydrated during such activities ensures safety. This dual perspective—heat as both waste and resource—highlights the ETC’s adaptability in meeting diverse physiological demands.
Electric Stoves: Energy Efficiency Myths and Real Electricity Costs
You may want to see also
Explore related products





![]()
Superoxide Radicals: Incomplete oxygen reduction can produce reactive oxygen species (ROS)
During the electron transport chain (ETC), oxygen typically serves as the final electron acceptor, forming water in a highly efficient process. However, this system is not infallible. Incomplete reduction of oxygen can occur, leading to the formation of superoxide radicals (O₂⁻), a type of reactive oxygen species (ROS). This happens when electrons prematurely leak from the ETC, particularly at Complexes I and III, reacting with molecular oxygen to produce these highly reactive molecules. While the body has evolved antioxidant defenses to neutralize ROS, an imbalance between production and detoxification can result in oxidative stress, damaging cellular components like DNA, proteins, and lipids.
Superoxide radicals are particularly insidious due to their ability to initiate a cascade of harmful reactions. For instance, superoxide can react with nitric oxide (NO) to form peroxynitrite (ONOO⁻), a potent oxidant that exacerbates cellular damage. Additionally, superoxide can be converted to other ROS, such as hydrogen peroxide (H₂O₂) and hydroxyl radicals (OH⁻), through enzymatic or metal-catalyzed reactions. These species are even more reactive and can cause irreversible damage to cells, contributing to aging, neurodegenerative diseases, and cancer. Understanding the mechanisms of superoxide formation is crucial for developing strategies to mitigate its harmful effects.
To combat superoxide-induced damage, the body employs enzymes like superoxide dismutase (SOD), which catalyzes the conversion of superoxide to hydrogen peroxide and oxygen. There are three isoforms of SOD: Cu/Zn-SOD in the cytoplasm, Mn-SOD in the mitochondria, and extracellular SOD (ECSOD). Supplementation with antioxidants such as vitamin C, vitamin E, and glutathione can also support the neutralization of ROS. For example, a daily intake of 15–30 mg of vitamin C and 15 mg of vitamin E is recommended for adults to maintain optimal antioxidant levels. However, excessive supplementation should be avoided, as it can disrupt the body’s natural redox balance.
Practical strategies to minimize superoxide production include lifestyle modifications that reduce oxidative stress. Regular physical activity, for instance, enhances mitochondrial efficiency and upregulates antioxidant defenses, but excessive exercise can paradoxically increase ROS production. A balanced diet rich in fruits, vegetables, and whole grains provides phytonutrients that bolster antioxidant capacity. For older adults, who are more susceptible to oxidative damage due to declining mitochondrial function, targeted interventions like caloric restriction or intermittent fasting may help reduce ROS generation. Monitoring biomarkers of oxidative stress, such as malondialdehyde (MDA) or 8-hydroxy-2'-deoxyguanosine (8-OHdG), can guide personalized approaches to managing superoxide-related risks.
In conclusion, superoxide radicals are a significant byproduct of incomplete oxygen reduction during the ETC, posing a threat to cellular integrity through their role in ROS production. By understanding their formation, leveraging enzymatic defenses, and adopting proactive lifestyle measures, individuals can mitigate the detrimental effects of oxidative stress. This knowledge not only highlights the importance of maintaining redox balance but also underscores the potential for targeted interventions to preserve health and prevent disease.
Digital X-Rays: Reducing Waste in Medical Imaging Practices
You may want to see also
Explore related products






![]()
Proton Gradient Collapse: Protons leak back into the matrix, reducing ATP synthesis efficiency
The electron transport chain (ETC) is a marvel of cellular efficiency, generating ATP through a proton gradient across the mitochondrial membrane. However, this system is not without its vulnerabilities. One critical issue arises when protons leak back into the mitochondrial matrix, a phenomenon known as proton gradient collapse. This leakage undermines the electrochemical potential necessary for ATP synthase to function optimally, directly reducing the efficiency of ATP synthesis. Understanding this process is essential for grasping the limitations of cellular energy production and exploring ways to mitigate its impact.
Proton leakage occurs primarily through uncoupling proteins (UCPs) and other channels in the inner mitochondrial membrane. These proteins act as pathways for protons to bypass ATP synthase, dissipating the gradient as heat instead of generating ATP. While this uncoupling can be beneficial in thermogenesis, such as in brown adipose tissue, it becomes detrimental when it occurs in excess. For instance, in skeletal muscle cells, uncontrolled proton leakage can reduce ATP yield by up to 20–30%, depending on metabolic demand and UCP activity. This inefficiency is particularly problematic during high-energy activities or in states of metabolic stress.
From a practical standpoint, minimizing proton gradient collapse is crucial for optimizing cellular energy output. One strategy involves modulating UCP activity through dietary interventions. For example, polyphenols like resveratrol and epigallocatechin gallate (EGCG) have been shown to inhibit UCPs, thereby preserving the proton gradient. Additionally, maintaining adequate levels of coenzyme Q10 (CoQ10), a key component of the ETC, can support efficient electron flow and reduce the likelihood of proton leakage. For adults over 40, supplementing with 100–200 mg of CoQ10 daily may help counteract age-related declines in mitochondrial function.
Comparatively, proton leakage is not solely a pathological concern but also a regulatory mechanism. In certain tissues, such as the brain, mild uncoupling can protect against oxidative stress by reducing the production of reactive oxygen species (ROS). However, this protective effect comes at the cost of ATP efficiency, highlighting the delicate balance between energy production and cellular defense. Researchers are now exploring pharmacological agents that selectively modulate UCPs to harness their benefits without compromising ATP synthesis, a promising avenue for treating metabolic disorders.
In conclusion, proton gradient collapse is a double-edged sword in the context of the electron transport chain. While it serves regulatory functions, its unchecked occurrence can significantly impair ATP synthesis. By understanding the mechanisms and consequences of proton leakage, individuals and researchers can adopt targeted strategies to enhance mitochondrial efficiency. Whether through dietary adjustments, supplementation, or emerging therapies, addressing this issue is vital for maintaining cellular energy homeostasis and overall metabolic health.
Do 1000cc Fuel Injectors Increase Gas Consumption? Find Out
You may want to see also
Frequently asked questions
The primary waste products of the electron transport chain are water (H₂O) and carbon dioxide (CO₂). Water is formed during the final step of oxidative phosphorylation when molecular oxygen (O₂) accepts electrons and combines with hydrogen ions (H⁺).
Water is produced when molecular oxygen (O₂) acts as the final electron acceptor in the electron transport chain. Electrons from NADH and FADH₂ are passed through the ETC complexes, and at the end, oxygen combines with these electrons and hydrogen ions (H⁺) from the matrix to form water (H₂O).
Carbon dioxide (CO₂) is not directly produced by the electron transport chain itself. Instead, it is generated during the earlier stages of cellular respiration, specifically in the citric acid cycle (Krebs cycle), where pyruvate derived from glycolysis is oxidized, releasing CO₂ as a byproduct. The ETC primarily produces water.































